Авторы Edward S. Bliss и Eliza Whiteside. Впервые опубликовано 09/2018 в журнале Frontiers in Physiology. Оригинал статьи доступен по ссылке.
Ожирение — это глобальная эпидемия, представляющая значительную социоэкономическую проблему для систем здравоохранения. Она особенно актуальна в западных странах, таких как Австралия, США, Великобритания и Канада. Ожирение развивается из-за нарушения баланса между потребляемой и расходуемой энергией, когда объем потребляемой энергии превышает объем расходуемой.
Современные неинвазивные методы лечения малоэффективны в борьбе с ожирением, демонстрируя, что это более многогранное и комплексное состояние, чем думали раньше. Это привело к росту исследований энергетического гомеостаза и к открытию системы двунаправленной связи, известной как кишечно-мозговая ось.
Кишечно-мозговая ось включает в себя различные нейрогуморальные компоненты, которые позволяют кишечнику и мозгу связываться друг с другом. Эта связь поддерживается с помощью локальных паракринных или/и эндокринных механизмов, включающих множество гастроинтестинальных пептидов, продуцируемых эндокринными клетками кишечника. Среди этих пептидов — глюканоподобный пептид-1 (GLP-1), холецистокинин (ССК), пептид YY (PYY), панкреатический полипептид (РР) и оксинтомодулин. Нервная система (в частности, энтеральная нервная система (ENS) и блуждающий нерв), тоже участвует в передаче информации внутри кишечно-мозговой оси. Появляются доказательства того, что кишечная микрофлора (комплексная экосистема, находящаяся в желудочно-кишечном тракте) может влиять на вес посредством взаимосвязанных механизмов (сбора энергии, передачи сигнала короткоцепочечными жирными кислотами, изменения поведения хозяина, контроля его насыщения и модулирования воспалительного ответа).
В этом обзоре будут рассмотрены кишечно-мозговая ось, микрофлора кишечника, связь между этими элементами и роль, которую каждый из них играет в активации и регулировании энергетического обмена и развитии ожирения.
Ожирение: растущая проблема
Ожирение — это одна из самых быстрорастущих эпидемий, вставших перед лицом международных систем здравоохранения, в особенности, развитых стран — Австралии, США, Великобритании и Канады. В 1970-е годы лишний вес и ожирение не были распространены, и меньше 15% австралийцев входили в эти категории (Hayes et al., 2017). К 1995 году доля людей с лишним весом и ожирением увеличилась приблизительно до 20% (Tolhurst et al., 2016; Hayes et al., 2017). Сейчас Австралия находится на первом месте в мире по распространенности лишнего веса и ожирения: 63,4% взрослых и 29,5% детей и подростков до 17 лет имеют избыточный вес (Grima and Dixon, 2013; Tolhurst et al., 2016). 44,5% взрослых и от 70,1 до 91,7% людей до 17 лет не получают необходимого минимума ежедневной физической активности, и примерно 40% ежедневно едят фаст-фуд. Такая избыточно калорийная диета, богатая насыщенными жирами, транс-жирами и простыми углеводами, получила название «западной». (Tolhurst at al., 2016). Причиной ожирения является отложение жиров из-за дисбаланса между потреблением и тратой энергии, когда потребление превышает трату. Исходя из этого упрощенного определения, ожирение — это следствие сложного комплекса генетических, диетических, социо-экономических, психологических и экологических факторов. Такое определение упрощает понимание ожирения и борьбу с ним как комплексным заболеванием (Moran and Shanahan, 2014; Bauer et al., 2016).
При ожирении наблюдается увеличение количества жировой ткани, развитие неспецифического воспаления, дисбактериоза и гормонального дисбаланса, повышение нейрогенного тонуса (Buhmann et al., 2014; Moran and Shanahan, 2014; Bauer et al., 2016). Эти факторы приводят к развитию коморбидностей (табл. 1), которые в свою очередь увеличивают частоту летальных исходов. Факторы, способствующие развитию ожирения, а также связанные с ними расходы (превышающие 8 млрд. долларов США в год только в Австралии) и провал неинвазивных методов лечения ожирения привели к увеличению числа исследований, направленных на улучшение подходов к снижению веса (Grima and Dixon, 2013; Buhmann et al., 2014). В настоящее время бариатрические операции, такие как желудочное шунтирование Roux-en-Y, лапароскопическая рукавная гастроэктомия и лапароскопическое бандажирование желудка, являются наиболее эффективными методами увеличения и поддержания потери веса в долгосрочной перспективе. Тем не менее, причина успеха бариатрических операций пока неясна. Предполагается, что ключом к пониманию этого явления могут стать изменения в системной и локальной концентрациях пептидов, продуцируемых кишечником, изменение ответа в местах их действия, а также изменения в работе блуждающего нерва и, соответственно, в сигналах, поступающих в головной мозг (Santo et al., 2016; Yavuz et al., 2017). Эта идея легла в основу большого числа исследований о взаимодействии между центральной нервной системой (ЦНС), желудочно-кишечным трактом (ЖКТ) (а с недавних пор – и микрофлорой кишечника) и энергетическим гомеостазом. В этом обзоре внимание сосредоточено на исследовании связи между осью «кишечник-мозг-микрофлора» и той ролью, которую каждый из звеньев оси играет и в активации, и в регуляции энергетического обмена, приводя к развитию ожирения.
| Система организма | Коморбидность | Ссылка |
|---|---|---|
| Сердечно-сосудистая | ИнсультИнфаркт миокарда
Стенокардия ИБС Сердечная недостаточность Артериальная гипертензия Тромбоз глубоких вен Эмболия легочной артерии Дислипидемия |
Wilson et al., 2002; Stein et al., 2011; Global Burden of Metabolic Risk Factors for Chronic Diseases Collaboration et al., 2014; Writing Group et al., 2014; Klovaite et al., 2015; Aune et al., 2016 |
| Желудочно-кишечная | НАЖБПЗаболевания желчного пузыря и поджелудочной железы
ГЭРБ Рак печени, пищевода, желчного пузыря; колоректальный рак |
Chen et al., 2012; Eslick, 2012; Stinton and Shaffer, 2012; DiBaise and Foxx-Orenstein, 2013 |
| Эндокринная | Инсулинонезависимый сахарный диабетГестационный сахарный диабет
Синдром поликистозных яичников |
Flegal et al., 2007; Arendas et al., 2008; Yang et al., 2008 |
| Мочеполовая | ХБПНефролитиаз
Рак почек, предстательной железы Недержание мочи Эректильная дисфункция Скрытый половой член |
Bump et al., 1992; Esposito et al., 2004; Ejerblad et al., 2006; Flegal et al., 2007; Polednak, 2008; Munkhaugen et al., 2009; Pestana et al., 2009; Stinton and Shaffer, 2012; Grima and Dixon, 2013 |
| Дыхательная | Синдром обструктивного апноэ снаСиндром ожирения-гиповентиляции
Астма ХОБЛ |
Guerra et al., 2002; Steuten et al., 2006; Eisner et al., 2007; O’Donnell et al., 2014 |
| Опорно-двигательная | ОстеоартритПоражения межпозвоночных дисков, поясничная боль
Поражения сухожилий, фасций и суставных хрящей Поражения стопы Нарушение мобильности |
Molenaar et al., 2008; Tukker et al., 2009; McAdams DeMarco et al., 2011; Grima and Dixon, 2013 |
| Репродуктивная | Нарушения менструального циклаОсложнения беременности (выкидыш, внутриутробная гибель плода)
Пороки развития Бесплодие Рак груди (постменопаузальный), эндометрия, яичников |
Bianchini et al., 2002; Arendas et al., 2008; Polednak, 2008; Grima and Dixon, 2013 |
| Нервная (в т.ч. высшие нервные функции) | ДеменцияДепрессия
Нарушения пищевого поведения Снижение качества жизни Психологическая стигматизация, снижение самооценки |
Beydoun et al., 2008; Molenaar et al., 2008; Grima and Dixon, 2013; Hilbert et al., 2014 |
| Покровная | Увеличение потоотделенияСнижение активности регенеративных процессов
Стрии Целлюлит Гиперпигментация Опрелость Лимфедема |
Löffler et al., 2002; Yosipovitch et al., 2004, 2007 |
| Иммунная | Поражения лимфоидной тканиНарушения развития лейкоцитов, их фенотипов и профиля активности
Снижение иммунитета Снижение эффективности вакцин Увеличение уровня провоспалительных маркеров (IL6, TNFα) |
Ghanim et al., 2004; Bremer et al., 2011; Kanneganti and Dixit, 2012; Sheridan et al., 2012 |
ЦНС, а в частности, головной мозг, выполняет сложную функцию анализа непрерывного потока информации об энергетическом состоянии организма, которую поставляют нервы и химические вещества. Эта информация используется для запуска подходящей для поддержания гомеостаза реакции. Сигналы варьируют, и ответы на них меняются в зависимости от типа потребляемой пищи. Хотя сначала пища попадает в полость рта, где сталкивается с оральной микрофлорой, ЖКТ остается основным местом сбора и переваривания пищи. Поэтому кишечник становится ответственным за генерацию большинства сигналов, идущих в ЦНС, о содержании и объеме поглощенной пищи. Таким образом создается сложную систему двунаправленной связи, которую называют кишечно-мозговой осью (Bauer et al., 2016; Gribble and Reimann, 2016).
Кишечно-мозговая ось: связи от кишечника к мозгу
Кишечно-мозговая ось — это комплексная нейрогуморальная связующая система, необходимая для поддержания метаболического гомеостаза. Ее элементами являются ЦНС, энтеральная нервная система (ЭНС), вегетативная нервная система (ВНС) и относящиеся к ней симпатические и парасимпатические ветви, нейроэндокринная и иммунная системы, а также кишечная микрофлора (Grenham et al., 2011). Связь внутри оси формируется посредством преобразования сенсорной информации в нейронные, гормональные и иммунные сигналы, передающиеся от ЦНС к кишечнику и обратно (Mayer et al., 2015). Хотя появляется все больше доказательств того, что изменения иммунной системы в кишечнике влияют на энергетический гомеостаз, большинство признанных эффектов оси является следствием работы нейронных и гормональных сигналов в кишечнике, т.к. ЖКТ обладает более, чем 500 миллионов нейронов и может продуцировать множество гормонов (Monje, 2017). Из-за большой степени иннервации ЖКТ питательные вещества перед всасыванием могут способствовать отправке в ЦНС сигналов об их калорийности и содержании макронутриентов через специфические сенсорные механизмы, расположенные по всему ЖКТ (Hamr et al., 2015). Эти сигналы в дальнейшим передаются в разные области головного мозга, такие как мозговой ствол и гипоталамус. Более высокая обработка сигналов в этих центрах позже инициирует ряд реакций, которые приводят как к острым, так и к хроническим изменениям в потреблении и расходе энергии. Это позволяет поддерживать метаболический гомеостаз до и после приема пищи (Buhmann et al., 2014).
Гормоны кишечника высвобождаются из эндокринных клеток кишечника (ЭКК). Они инициируют большинство сигналов внутри кишечно-мозговой оси в ответ на воздействие питательных веществ перед их всасыванием. Эти клетки находятся в эпителии ЖКТ, и апикальные мембраны многих из них покрыты микроворсинками, непосредственно контактирующими с содержимым просвета ЖКТ (Gribble and Reimann, 2016). Схема работы ЭКК представлена ниже (рис. 1).
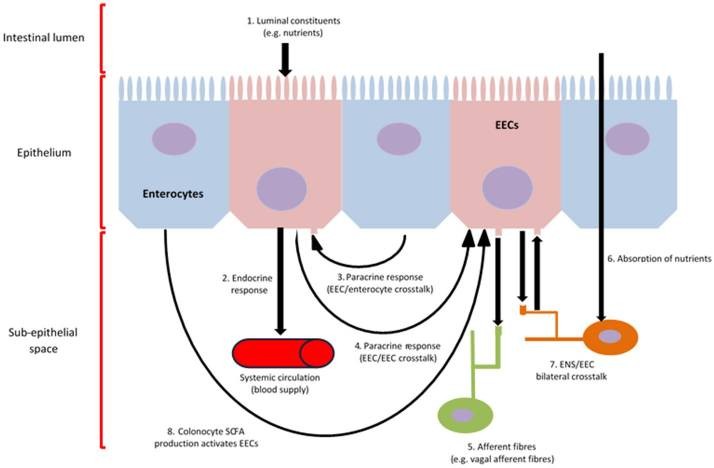
Рис. 1. Функции и связи эндокринных клеток кишечника. Внутриклеточный метаболизм и активация хеморецепторов, расположенных на апикальной клеточной мембране ЭКК, приводят к притоку кальция в клетку, который индуцирует синтез и высвобождение гормонов кишечника в субэпителиальное пространство (1, 4) (Psichas et al., 2015). Различные гормоны, полученные из кишечника, синтезируются и секретируются в ответ на вещества в просвете и систематически высвобождаются из ЭКК, чтобы воздействовать на различные ткани организма, такие как мозг, через метаболический, местный, паракринный (3) и/или эндокринный (2) механизмы, а также активировать афферентные нейроны, иннервирующие стенку ЖКТ (5, 6, 7, 8) (Psichas et al., 2015). Кроме того, нарушения в связи ЭКК/ЭНС могут быть результатом прямого всасывания питательных веществ в кишечнике (7). Производство КЖК микрофлорой, которые впоследствии могут быть использованы колоноцитами в качестве источника энергии, может активировать ЭКК, что будет способствовать активации кишечно-мозговой активности (8).
Основной объем переваривания и всасывания происходит в желудке и в тонком кишечнике, и поэтому эти органы сильно иннервированы: они являются первичными участками, ответственными за улавливание содержания питательных веществ в поступающем пищевом комке. За их иннервацию отвечают блуждающий и внутренностные нервы (Bauer et al., 2016). Здесь количество афферентных волокон превосходит количество эфферентных волокон, что указывает на фундаментальную роль нейронной от кишечника к мозгу (Prechtl and Powley, 1990; Berthoud et al., 1995). Волокна блуждающего нерва подходят к собственным пластинкам кишечных ворсинок, заканчиваются на базолатеральной мембране эндокринных клеток кишечника и экспрессируют рецепторы к гормонам кишечника, таким как грелин, лептин, холецистокинин, глюканоподобный пептид-1 и пептид YY (PYY). Активация рецептора приводит к последующей нейронной активации (Dockray, 2013). Кроме того, нейроны ЭНС, обладающие рецепторами для разных гастроинтестинальных гормонов, могут косвенно активировать волокна блуждающего и внутренностных нервов (Almato et al., 2010; Richards et al., 2014). В то время, как ЭНС контролирует кишечную функцию локально через рефлекторные действия, нельзя не отметить ее роль в передаче сигналов о питательных веществах афферентным волокнам блуждающего нерва, что способствует работе кишечно-мозговой оси (Costa et al., 2000; Sayegh et al., 2004). Нейроны ЭНС расположены проксимально как относительно ЭКК, так и относительно окончаний афферентных нервов; они стимулируются введением питательных веществ в кишечник и активируют афферентные волокна блуждающего нерва в кишечнике (Sayegh et al., 2004, Ritter, 2011). Хотя точный механизм работы ЭНС не выяснен, а мнение о том, что ЭНС работает независимо от ЦНС, все еще превалирует, из этих исследований очевидно, что нейронный сигнал в кишечно-мозговой оси возникает из-за гормональной секреции кишечника, вызванной поступлением питательных веществ.
При приеме пищи сенсорная информация поступает из афферентных волокон блуждающего нерва и/или соматосенсорной системы в ядро одиночного пути (nucleus tractus solitarii — NTS). Более конкретно, афферентные волокна блуждающего нерва сходятся в NTC дорсального вагального комплекса внутри мозгового ствола, а соматосенторные афферентные волокна образуют синапсы с нейронами пограничной зоны заднего рога спинного мозга, которые затем проецируются в NTS (Zittel et al., 1994, Schwartz et al., 2000). NTS, в свою очередь, собирает и доставляет сигналы из ЖКТ в гипоталамус (Craig, 1996; Schwartz et al., 2000). При использовании c-Fos—a-маркера, выявляющего повышенную активность нейронов (Zittel et al., 1994), был обнаружен рост его экспрессии в NTS во время всасывания пищи в ЖКТ. В то же время, введение высокой дозы нейротоксина капсаицина уменьшало экспрессию c-Fos—a-маркера и блокировало связь кишечника с мозгом через блуждающий нерв (Mönnikes et al., 1997). Кроме того, в исследовании Campos et al. (2013) утверждается, что нейроны NТС стимулировались афферентами блуждающего нерва путем активации рецепторов N-метил-D-аспартата (NMDA-рецепторы) в афферентных окончаниях. Впоследствии это приводило к высвобождению нейромедиаторов путем фосфорилирования внеклеточных сигнальных киназ 1/2 и синапсина 1. Babic et al. (2009) установили, что другие нейроны NTC активируются через афференты блуждающего нерва, стимулирующие \проопиомеланокортиновые (ПОМК) и катехоламинергические нейроны. Считалось, что эти нейроны NTC участвуют в сообщении о насыщении через сигнальные меланокортиновые рецепторы внутри гипоталамуса (рис. 2). Интересно, что недостаточность меланокортинового рецептора 4 типа способствует развитию ожирения (Farooqi et al., 2003). Более точные исследования механизмов взаимодействий различных рецепторов и нейронов могут привести к более глубокому пониманию того, как нейроны NTC способствуют энергетическому гомеостазу. Кроме того, результаты, указывающие на стимуляцию POMC и катехоламинергических нейронов через волокна блуждающего нерва, а также на наличие NMDA-рецепторов в NTS, могут помочь в понимании связи между потреблением пищи и модификациями поведения, учитывая, что эти нейроны высвобождают нейромедиаторы, связанные с возбуждением, мотивацией и эмоциями (к примеру, дофамин).
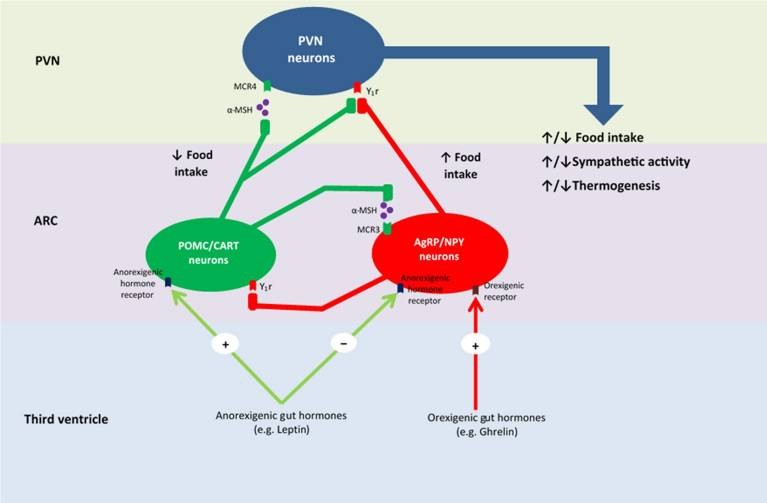
Рис. 2. Предлагаемый механизм энергетического гомеостаза внутри гипоталамуса. PVN — паравентрикулярное ядро; ARC — дугообразное ядро; MCR4 — рецептор меланокортина 4; α-MSH — α- меланокортин-стимулирующий гормон; MCR3 — рецептор меланокортина 3; Y1r — рецептор нейропептида Y типа 1; POMC — про-опиомеланокортин; CART — транскрипт, регулируемый кокаином и амфетамином; NPY — нейропептид Y; AgRP — агути-подобный белок.
Нейроны NTS заканчиваются в нескольких более высоких центрах головного мозга, включая систему меланокортина, в которую входит гипоталамус (Suzuki et al., 2012). Гипоталамус выполняет фундаментальную роль интеграции периферических гуморальных сигналов, которые передают информацию о потреблении питательных веществ и энергозатратах, а также информацию из NТС и других областей мозга (Bauer et al., 2016). Особую роль в пищевом поведении играют дугообразное, паравентрикулярное, вентромедиальное и дорсомедиальные ядра гипоталамуса и латеральный отдел гипоталамуса (Сone et al., 2001; Suzuki et al., 2012). Эти зоны объединены сетью, регулирющей энергетический гомеостаз. Большинство исследований было сосредоточено на дугообразном ядре и его участии в поддержании энергетического гомеостаза (Suzuki et al., 2012; Buhmann et al., 2014). Следовательно, необходимо изучение роли оставшихся зон гипоталамуса в пищевом поведении и регулировании энергетического гомеостаза. Впрочем, ясно, что чувствительность к питательным веществам появляется в ЖКТ и вызывает множество нейронных и/или гуморальных ответов, которые вносят основной вклад в работу кишечно-мозговой оси, участвующей в регуляции энергетического баланса.
Дугообразное ядро отвечает на периферические и центральные сигналы об аппетите через тщательно регулируемое выделение нейромедиаторов из двух групп нейронов: проопиомеланокортиновые (ПОМК) нейроны и нейроны, связанные с агути-подобным (AGRP) пептидом. AGRP-нейроны, расположенные в в медиальной части дугообразного ядра, выбрасывают ингибирующие нейромедиаторы-AGRP и нейропептид Y (NPY) (Сone et al., 2001; Suzuki et al., 2012). Эти нейромедиаторы стимулируют голод и аппетит, а также уменьшают расход энергии. Это увеличивает потребление пищи и способствует набору веса (Dryden et al., 1995; Ollmann et al., 1997; Enriori et al., 2007). ПОМК-нейроны в латеральной части дугообразного ядра выбрасывают ПОМК, который стимулирует выделение альфа-меланоцит-стимулирующего гормона (α-MSH) и СART-пептида (cocaine-amphetamine-regulated transcript) (Suzuki et al., 2012). Эти нейромедиаторы являются антагонистами AGRP и NPY, уменьшают аппетит и чувство голода, что снижает потребление пищи, а также стимулируют увеличение расхода энергии. Все эти факторы способствуют потере веса (Cowley et al., 2001; Nakhate et al., 2011). Энергетический гомеостаз зависит от сбалансированной работы этих двух групп нейронов.
Регуляторные механизмы гипоталамуса усложняются за счет того, что на дугообразное ядро напрямую влияют гастроинтестинальные гормоны, количество которых увеличивается после приема пищи (van der Kooy, 1984). Дугообразное ядро и ядро одиночного пути (NТС) находятся в зоне мозга, где гематоэнцефалический барьер проницаем. Эта зона называется самым задним полем (area postrema — AP). Повреждение АP и ваготомия уменьшают действие гастроинтестинальных гормонов, указывая на то, что высвобождение этих гормонов в системный кровоток напрямую влияет на эту зону (van der Kooy, 1984; Date et al., 2002). Batterham et al. (2002) проиллюстрировали это с помощью введения инъекции PYY в основной кровоток. PYY связывался с рецептором Y2, который Batterham et al. (2002) локализовали в ARC, обнаружив увеличение иммунореактивности c-FOS. Кроме того, Seeley et al. (1994) установили, что у децеребрированных крыс с сохранным мозговым стволом происходило подавление пищевого потребления и увеличение энергозатрат в ответ на всасывание питательных веществ в кишечнике. Это раннее исследование, проведенное в 1990-х годах, легло в основу дальнейших открытий о том, что сигналы из кишечника систематически попадают в NТС через проницаемый гематоэнцефалический барьер (Bauer et al., 2016). Учитывая отсутствие у этих крыс головного мозга, интересно, что у них были увеличены затраты энергии (Seeley et al., 1994). Это подразумевает, что мозговой ствол регулирует механизм затраты энергии в качестве проекционной зоны. Требуются более глубокие исследования о связи работы кишечника и мозгового ствола. Они могут помочь в обосновании связи между потреблением пищи и упражнениями и роли, которую каждый из этих факторов играет по отношению к энергетическому гомеостазу.
Гастроинтестинальные гормоны и роль кишечно-мозговой оси в энергетическом гомеостазе
Обсудив связи с ЦНС, проанализируем роль желудка в потреблении питательных веществ. Желудок — один из первых органов, откуда поступают сигналы в меланокортиновую систему. Когда пищевой комок попадает в ЖКТ, желудок растягивается, сигнализируя мозгу о необходимости прекращении приема пищи. Поступление пищи из желудка в двенадцатиперстную кишку происходит при расслаблении пилорического сфинктера. Как только питательные вещества попадают в двенадцатиперстную кишку, скорость опорожнения желудка уменьшается, что увеличивает его растяжение и ограничивает количество потребляемой пищи.
Скорость опорожнения желудка снижается при активации блуждающего нерва и высвобождении гормонов кишечника, таких как CCK, PYY и GLP1 (Cooke and Clark, 1976; Talsania et al., 2005; Suzuki et al., 2012). Этот механизм обратной связи был продемонстрирован в исследовании Davis and Smith (1990), которые установили, что потребление пищи при повторном кормлении у крыс уменьшается в течение 6 минут после первого кормления, предотвращая чрезмерное потребление. Phillips and Powley (Phillips and Powley, 1996) продемонстрировали на крысах, использyя физиологический раствор вместо пищевых продуктов, что именно растяжение желудка, а не содержание питательных веществ в пищевом комке, вызывает сигнал обратной связи, прекращающий чрезмерное потребление, и что для его развития требуется менее чем 3 минуты. Эти исследования показывают, что иннервирующие желудок нейроны экспрессируют механорецепторы, которые активируются растяжением желудка, способствуют передаче ограниченной информации в мозг и участвуют в регулировании питания и долгосрочного энергетического гомеостаза (Bauer et al. 2016). Поступают также новые данные о содержании в желудке вкусовых рецепторов. Они могут противоречить предыдущим исследованиям о природе сенсорной информации, передаваемой в ЦНС, и о том, как ЦНС интегрирует эту информацию и передает ее остальной части кишечника (Young et al., 2009; Depoortere, 2014). Классические исследования, впрочем, демонстрируют, что ложный прием пищи (процесс, при котором пищевые продукты, поступающие в желудок, не попадают в тонкую кишку и попадают непосредственно в толстую кишку, или удаляются непосредственно из желудка) замедляется с помощью инфузии кишечного питательного вещества, и тем самым доказывают, что питательные вещества в кишечнике могут подавлять потребление пищи независимо от опорожнения желудка и передачи информации на кишечно-мозговую ось с помощью нейрогуморальных механизмов (Gibbs et al., 1981; Reidelberger et al., 1983).
Далее будет рассмотрена роль гормонов кишечника в качестве контролеров приема пищи в отношении энергетического гомеостаза. Поскольку эффекты глюкагона, инсулина, лептина и грелина обсуждаются ниже, они не будут пересмотрены в этом обзоре, и их функции суммируются в табл. 2 (Sakata and Sakai, 2010; Dimitriadis et al., 2011; Jones et al., 2012; Pan et al., 2014).
| Гормон | Тип энтероцита | Основная локализация | Функция | Системная концентрация
(при ожирении) |
Системная концентрация
(после бариатрической операции) |
Ссылки |
|---|---|---|---|---|---|---|
| CCK | I-клетки | Двенадцатиперстная кишка, тощая кишка | Опорожнение желудка ↓Поджелудочная секреция ↑
Чувство насыщения ↑ |
Неясно | Без изменений | Gibbs et al., 1973; van der Kooy, 1984; Calingasan et al., 1992; Lieverse et al., 1995; Kopin et al., 1999; Di Francesco et al., 2005; Cheung et al., 2009; Rasmussen et al., 2012; Duca and Yue, 2014; Dixon et al., 2015; Münzberg et al., 2015 |
| GLP1 | L-клетки | Подвздошная кишка | Секреция инсулина ↑Пролиферация β-клеток ↑
Экспрессия генов β-клеток ↑ Чувство насыщения ↑ Секреция желудочной кислоты ↓ Опорожнение желудка ↓ Апоптоз ↓ |
↓ | ↑ | Elliott et al., 1993; Deacon et al., 1995; Larsen et al., 1997; Yamato et al., 1997; Edwards et al., 1999; Villanueva-Peñacarrillo et al., 2001; Vilsbøll et al., 2003; Baggio et al., 2004; Abbott et al., 2005; Adam and Westerterp-Plantenga, 2005; Cani et al., 2005; Holst, 2007; Li et al., 2009; Pocai et al., 2009; Rüttimann et al., 2009; Suzuki et al., 2012; Wang et al., 2012; Zhang and Ritter, 2012; Mumphrey et al., 2013; Ohlsson et al., 2014; Richards et al., 2014; Dixon et al., 2015; Kuhre et al., 2015; Münzberg et al., 2015; Svendsen et al., 2015; Graaf et al., 2016 |
| PYY | L-клетки | Подвздошная кишка | Секреция желудочной кислоты ↓Поджелудочная и кишечная секреция ↓
Моторика ЖКТ ↓ Чувство насыщения ↑ |
↓ | ↑ | Adrian et al., 1985; Fu-Cheng et al., 1997; Batterham et al., 2002, 2003a, 2006; Abbott et al., 2005; Di Francesco et al., 2005; Koda et al., 2005; Korner et al., 2005; Stock et al., 2005; Talsania et al., 2005; le Roux et al., 2006; Oesch et al., 2006; Pyarokhil et al., 2012; El-Salhy et al., 2013; Dixon et al., 2015; Münzberg et al., 2015 |
| PP | F-клетки | Поджелудочная железа | Опорожнение желудка ↓Уровень лептина (белая жировая ткань) ↓
Чувство насыщения ↑ |
↓ | Без изменений | Adrian et al., 1976, 1985; Schwartz et al., 1978; Lassmann et al., 1980; Clark et al., 1984; Berntson et al., 1993; Parker and Herzog, 1999; Batterham et al., 2003b; Balasubramaniam et al., 2006; Lin et al., 2009; Habib et al., 2012; Kohno and Yada, 2012; Pyarokhil et al., 2012; Dixon et al., 2015; Khandekar et al., 2015 |
| Оксинтомодулин | L-клетки | Подвздошная кишка | Опорожнение желудка ↓Секреция желудочной кислоты ↓
Уровень глюкозы крови ↓ Секреция инсулина ↑ Потребление энергии ↑ Чувство насыщения ↑ |
↓ | Неясно | Anini et al., 1999; Dakin et al., 2002, 2004; Cohen et al., 2003; Baggio et al., 2004; Wynne et al., 2005, 2006; Maida et al., 2008; Druce et al., 2009; Pocai et al., 2009; Habib et al., 2012; Pocai, 2014; Meek et al., 2016 |
| Серотонин | Энтерохромаффинные клетки | Различные области | Сокращения кишечника ↑Моторика кишечника ↑
Чувство тошноты ↑/↓ Синтез оксида азота ↑ Интенсивность всасывания ↑ Влияет на различные неврологические и психологические процессы |
Неясно | Неясно | Halford et al., 1997; Savastano et al., 2007; Lam et al., 2008; Bertrand and Bertrand, 2010; Bertrand et al., 2011; Daly et al., 2011; Halford and Harrold, 2012; Suzuki et al., 2012; Mumphrey et al., 2013; Crane et al., 2015; Dixon et al., 2015; Hall, 2015; Münzberg et al., 2015; Stiedl et al., 2015; Palamiuc et al., 2017 |
| GIP | К-клетки | Двенадцатиперстная кишка, тощая кишка | Секреция инсулина ↑Пролиферация β-клеток ↑
Чувство насыщения ↑ Активность липопротеин липазы ↑ Отложение жировой ткани ↑ Синтез жирных кислот ↑ Синтез костной ткани ↑ Апоптоз ↓ |
↑ | Неясно | Elliott et al., 1993; Meier et al., 2001; Stock et al., 2005; Laferrère et al., 2008; Mentlein, 2009; Mieczkowska et al., 2013; Cho et al., 2015; Meek et al., 2016 |
| Гастрины | G-клетки | Желудок | Секреция желудочной кислоты ↑Созревание париетальных клеток ↑
Фундальный рост ↑ Секреция зимогена ↑ Сокращения желудка ↑ Чувство насыщения ↑ Опорожнение желудка ↓ Поджелудочная секреция ↑ Опорожнение желчного пузыря ↑ |
Неясно | Неясно | Valenzuela et al., 1976; Kazumori et al., 2001; Nørsett et al., 2008, 2011; Dimaline and Varro, 2014; Dixon et al., 2015; Hall, 2015; Münzberg et al., 2015 |
| Гистамин | Энтерохромаффинные клетки | Различные области | Влияет на различные неврологические и иммунологические процессыВазодилатация ↑
Секреция желудочной кислоты ↑ Выброс серотонина ↑/↓ |
Неясно | Неясно | Powell, 1976; Jutel et al., 2001; Parmentier et al., 2002; Li et al., 2003; Panula et al., 2015; Yuan et al., 2017 |
| Нейротензин | N-клетки | Тощая кишка | Влияет на различные неврологические и психологические процессыЧувство насыщения ↑
Опорожнение желудка ↓ Секреция желудочной кислоты ↓ Расход энергии ↓ Абсорбция жира ↑ Сокращения гладкой мускулатуры ↑/↓ |
Неясно | Неясно | Blackburn et al., 1980; Katz et al., 2004; Fawaz et al., 2009; Feifel et al., 2010; Kalafatakis and Triantafyllou, 2011; Dirksen et al., 2013; Grunddal et al., 2016; Li et al., 2016 |
| Секретин | S-клетки | Двенадцатиперстная кишка, тощая кишка | Секреция желудочной кислоты ↓Синтез бикарбоната ↑
Синтез желчи ↑ Сокращения желчного пузыря ↑ pH ↑ Секреция воды ↑ Секреция инсулина ↑ |
↓ | ↑ | Boyer and Bloomer, 1974; Gray et al., 1988; Chu et al., 2007; Cheng et al., 2011; Wang et al., 2015; Meek et al., 2016 |
| Глюкагон | α-клетки | Поджелудочная железа | Уровень глюкозы крови ↑Расход энергии ↑ | ↑ | ↓ | Billington et al., 1991; Dicker et al., 1998; Korner et al., 2006; Swarbrick et al., 2008; Ortega et al., 2011; Jones et al., 2012; Dixon et al., 2015; Münzberg et al., 2015 |
| Амилин | β-клетки | Поджелудочная железа | Опорожнение желудка ↓Секреция желудочной кислоты ↓
Увеличение постпрандидального уровня глюкозы ↓ |
↑ | ↓ | Higham et al., 2000; Ratner et al., 2004; Dixon et al., 2015; Hay et al., 2015; Münzberg et al., 2015 |
| Грелин | Α-подобные клетки | Желудок, двенадцатиперстная кишка | Секреция гормона роста ↑Моторика ЖКТ ↑
Вазодилатация ↑ Сократимость миокарда ↑ Чувство голода ↑ |
↓ | Неясно | Kojima et al., 1999; Bedendi et al., 2003; Korner et al., 2005; Stock et al., 2005; Sakata and Sakai, 2010; Mumphrey et al., 2013; Dixon et al., 2015; Münzberg et al., 2015; Yavuz et al., 2017 |
| Инсулин | β-клетки | Поджелудочная железа | Абсорбция ↑Синтез гликогена ↑
Уровень глюкозы крови ↓ Синтез липидов ↑ Липолиз ↓ Протеолиз ↓ Аутофагия ↓ |
↑ | ↓ | Korner et al., 2006; Shak et al., 2008; Swarbrick et al., 2008; Dimitriadis et al., 2011; Ortega et al., 2011; Kleinridders et al., 2014; Dixon et al., 2015; Pankov, 2016 |
| Лептин | Адипоциты (не являются энтероцитами) | Белая жировая ткань | Чувство голода ↓Секреция α-MSH ↑
Расход энергии ↑ Действие NPY и AgRP ↓ Экспрессия эндоканнабиноидов ↓ |
↑ | ↓ | Elias et al., 1999; Cowley et al., 2001; Ravinet Trillou et al., 2004; Korner et al., 2005; Enriori et al., 2007; Liu et al., 2011; Pan et al., 2014; Dixon et al., 2015; Münzberg et al., 2015; Yavuz et al., 2017 |
Холецистокинин (CCK)
СК – первый желудочно-кишечный гормон, который связали с контролем аппетита. Он секретируется кишечными эндокринными клетками в двенадцатиперстной кишке и тощей кишке после приема пищи (Gibbs et al., 1973). Его высвобождение стимулируется потреблением жиров и белков, а его концентрация увеличивается в течение 15 минут после приема пищи (Lieverse et al., 1995; Buhmann et al., 2014). CCK обладает коротким периодом полураспада (в течение нескольких минут) и, следовательно, имеет ограниченное время, чтобы воздействовать на рецепторы CCK-1 и CCK-2, расположенные в тканях ЖКТ и ЦНС, например, в блуждающем нерве, NTS и гипоталамусе (Buhmann et al., 2014; Lo et al., 2014). CCK увеличивает двигательную активность ЖКТ и желчного пузыря и их секрецию, в дополнение к тому, что играет значительную роль в иницииации контроля кишечно-мозговой осью потребления пищи, расхода энергии и использования глюкозы (Cheung et al., 2009; Suzuki et al., 2012). Периферическое введение CCK, проведенное в ходе исследований на животных, показывает его дозозависимый эффект на регуляцию потребления пищи, а введение антагонистов рецептора CCK-1 в сочетании с потреблением жирных кислот и белков препятствует стимуляции блуждающих афферентов, выстилающих тонкий кишечник, и развитию регуляторного влияния CCK на потребление пищи (Calingasan et al., 1992; Cox et al., 1996; Duca and Yue, 2014). Следовательно, CCK является специфическим медиатором насыщения жирами и белками. Повторные дозы CCK, попадающие в общий кровоток, и спонтанное всасывание CCK во время кормления уменьшают количество потребляемой пищи. Впрочем, со временем повышается толерантность организма к CCK, и увеличивается количество и частота приема пищи внутрь (Kopin et al., 1999; Buhmann et al., 2014). Этот десенсибилизирующий эффект может объяснить неудачные попытки использовать CCK-производные гормоны, такие как GI 181771X, для потери веса (Castillo et al., 2004; Kim et al., 2011).
ССК оказывает влияние на обмен глюкозы — он увеличивает активацию блуждающего нерва, который, в свою очередь, индуцирует NDMA-нейроны NTS для проведения сигналов в печень для снижения глюконеогенеза (Rasmussen et al., 2012). Когда крыс переводят на диету с высоким содержанием жиров и углеводов, они развивают CCK-резистентность в ответ на повышение дозы ССК (Daly et al., 2011). Точные механизмы глюкорегуляторного действия CCK и развития CCK-резистентности остаются неясными. Однако эти данные могут дать объяснение, почему CCK-производные индуцируют панкреатит и способствуют замедленному гликолизу. Также более детального изучения требует процесс влияния ССК на другие органы, например, на печень.
Глюкагоноподобный пептид-1 (GLP1)
GLP1 представляет собой нейропептид, продуцируемый преимущественно эндокринными клетками подвздошной кишки и толстой кишки в ответ на потребление углеводов, липидов и/или белков (Elliott et al., 1993; Adam and Westerterp-Plantenga, 2005). Он синтезируется посредством посттрансляционного процессинга препроглюкагона в ЦНС и ЖКТ, а затем активирует рецепторы GLP1 (тип GPCR, экспрессируемый в ЦНС, ЖКТ и поджелудочной железе) (Larsen et al., 1997; Yamato et al., 1997). Введение GLP1 стимулирует центры сытости в мозге, в частности, дугообразные, паравентрикулярное ядра, NTS и AP, для снижения чувства голода (Larsen et al., 1997; Abbott et al., 2005). Следовательно, GLP1 считается ключевым фактором, приводящим к насыщению. Он синтезируется и высвобождается в течение 15 минут после приема пищи. Это представляет особый интерес, если учитывать, что кишечные L-клетки (эндокринные клетки, которые его выделяют) расположены дистально в подвздошной кишке (Elliott et al., 1993; Bauer et al., 2016). Таким образом, с учетом их участия в начальной стадии пищеварения, высвобождение GLP1 может быть рефлекторным ответом с участием волокон блуждающего нерва, расположенных в пределах двенадцатиперстной кишки. Хотя эта гипотеза еще не подтверждена, недавние исследования продемонстрировали наличие GLP1-секретирующих эндокринных клеток в пределах двенадцатиперстной кишки. Это демонстрирует, что высвобождение GLP1 может происходить в два этапа или в ответ на этот гипотетический рефлекс (Svendsen et al., 2015). Более сложные исследования, включая исследования с вливанием питательных веществ в участки тонкой кишки, необходимы для того, чтобы определить, какой именно участок/ки секретирует GLP1. Пока ясно, что его высвобождение влияет на потребление энергии, чувствительно ко всем типам поступающих макромолекул и вызывает чувство сытости.
GLP1 является мощным инкретином (гормоном, регулирующим уровень глюкозы в крови), который стимулирует рецептор GLP1 панкреатических β-клеток для высвобождения инсулина (Buhmann et al., 2014). Кроме того, повышенный уровень GLP1 увеличивает экспрессию гена фактора 1 промотора инсулина β-клеток поджелудочной железы, способствует их развитию и препятствует их апоптозу. В свою очередь, это активизирует процессы утилизации глюкозы в организме (Villanueva-Peñacarrillo et al., 2001; Suzuki et al., 2012). Наконец, GLP1 уменьшает скорость опорожнения желудка в двенадцатиперстную кишку и препятствует секреции желудочного сока, что, в свою очередь, увеличивает желудочное растяжение, ограничивает чрезмерное потребление пищи, усиливает насыщение и оказывает положительное влияние на энергетический гомеостаз (Edwards et al., 1999).
Хотя GLP1 оказывает мощный эффект, его действие непродолжительно, поскольку GLP1 уязвим для быстрой деградации и инактивации посредством катализа дипептидилпептидазой IV (DPPIV) (Deacon et al., 1995; Holst, 2007). Только 10% GLP1, получаемого из кишечника, достигает системного кровообращения. Это указывает на то, что он передает свои эффекты паракринным образом (Vilsbøll et al., 2003; Holst, 2007; Kuhre et al., 2015). Кроме того, периферическое введение GLP1 в сочетании с удалением блуждающего нерва препятствует действию GLP1, в то время как внутрисосудистая инфузия GLP1 продолжает оказывать эффект даже после ваготомии и/или введения высокой дозы нейротоксина капсаицина (Rüttimann et al., 2009; Zhang and Ritter, 2012). Это указывает на то, что рецептор GLP1 расположен в мозге, и что GLP1 может функционировать и на более высоком уровне в качестве нейротрансмиттера, хотя его точные функции на этом уровне еще только предстоит определить. GLP1 стимулирует определенные области мозга, такие как мозговой ствол, для увеличения моторного ответа и/или термогенеза, но все еще необходимы дальнейшие исследования для определения точных механизмов его участия в регуляции энергетического гомеостаза и пищевого поведения. (Li et al., 2009; Graaf et al., 2016). Ohlsson et al. (Ohlsson et al., 2014) показали, что концентрация GLP1 в лимфе быстро растет после приема пищи и остается повышенной в течение большего промежутка времени, так как концентрация DPPIV в лимфе ниже, чем в системном кровотоке. Это предоставляет еще один возможный механизм центрального и периферического действия GLP1, а также возможное объяснение его роли в иммунной сигнализации и развитии воспаления, связанного с ожирением. Как бы то ни было, будущие исследования, направленные на расширение понимания функции GLP1 и/или имитацию его действия посредством агонистов рецепторов GLP1 (к примеру, экзенатида) могут предоставить новые сведения о механизмах насыщения и энергетического гомеостаза.
Пептид YY3−36 (PYY)
PYY — небольшой пептид, принадлежащий к семейству панкреатических пептидов, и, подобно GLP1, секретирующийся кишечными L-клетками после приема пищи (Batterham et al., 2002, 2006). Он выделяется в ответ на выявление кишечником поступивших питательных веществ в объеме, соответствующем количеству потребляемой энергии (Oesch et al., 2006). PYY секретируется 15 минут после приема пищи, и похож на GLP1 по механизмам чувствительности к пище, находящейся в двенадцатиперстной кишке, регулирования активности блуждающего нерва и химических реакций (Fu-Cheng et al., 1997). В отличие от CCK и GLP1, концентрации которых быстро снижаются, PYY сохраняет повышенную концентрацию в течение нескольких часов после приема пищи (Batterham et al., 2003a). Следовательно, эффекты PYY могут быть пролонгированы и более ярко выражены по сравнению с CCK и GLP1.
PYY присутствует на протяжении всего ЖКТ, от пищевода до прямой кишки (Adrian et al., 1985). PYY связывается с рецептором Y2 и уменьшает потребление пищи. Грызуны, лишенные этого рецептора или с недостаточностью PYY становятся прожорливыми и, следовательно, набирают вес (Batterham et al., 2006; le Roux et al., 2006). Кроме того, PYY вызывает активацию нейронов NTS и ПOMК в дугообразном ядре и при периферическом, и при центральном введении, что указывает на наличие рецепторов Y2 в блуждающем нерве, в NTS и в дугообразном ядре (Batterham et al., 2002, 2006; Koda et al., 2005; le Roux et al., 2006). PYY оказывает свое действие, ингибируя нейроны NPY в дугообразном ядре. Поскольку эти нейроны в изобилии экспрессируют Y2-рецепторы, их активация препятствует действию повышающего аппетит NPY (Dryden et al., 1995; Broberger et al., 1997). Таким образом, PYY может играть ключевую роль в энергетическом гомеостазе, эндокринно регулируя потребление пищи и подавляя чрезмерное потребление посредством активации нейронов ПОМК и ингибирования NPY в меланокортиновой системе (Bauer et al., 2016). Кроме того, некоторые исследования показывают, что пациенты с ожирением обладают более низкими концентрациями PYY после приема пищи, в то время как в других исследованиях было высказано предположение о большой разнице концентраций PPY натощак у людей с нормальной массой тела и у людей с ожирением (Batterham et al., 2003a; Korner et al., 2005; Stock et al., 2005). Увеличенные концентрации PYY связаны с желудочно-кишечными заболеваниями (ВЗК и хронический деструктивный панкреатит), в дополнение к длительной потере аппетита (El-Salhy et al., 2013). Кроме того, устойчивые концентрации PYY и CCK у пожилых людей сопровождаются задержкой опорожнения желудка и сниженной сократимостью желчного пузыря (Di Francesco et al., 2005; Buhmann et al., 2014). Механизм, лежащий в основе этого явления, остается неясным. Однако повышенные концентрации PYY, которые способствуют долговременному насыщению и, следовательно, снижению потребления энергии, могут быть связаны у пожилых людей с недоеданием (Di Francesco et al., 2005; Buhmann et al., 2014). До того, как агонисты рецептора Y2 и/или производные PYY могут быть использованы в качестве эффективного лечения ожирения, необходимы дальнейшие исследования для определения долгосрочных эффектов их повышенных концентраций.
Панкреатический полипептид
Панкреатический полипептид (РР) относится к семейству панкреатических пептидов и секретируется специализированными F-клетками в островках Лангерганса (Khandekar et al., 2015). Его выделение, как и PYY, пропорционально потреблению калорий – продукты с высоким содержанием жира стимулируют его секрецию (Guyenet and Schwartz, 2012). Он выделяется системно до и во время приема пищи, механизм секреции похож на механизм секреции GLP1 и PYY в связи с чувствительностью к пище в двенадцатиперстной кишке, химическим реакциям и активации волокон блуждающего нерва (Schwartz et al., 1978; Khandekar et al., 2015). Было продемонстрировано, что его высокая концентрация поддерживается вплоть до 6 ч после приема пищи, что предполагает наличие эндокринного действия (Adrian et al., 1976). Кроме того, РР выделяется в толстой кишке и прямой кишке быка, где он действует как гормон внешней секреции (Pyarokhil et al., 2012). Наличие этой функции у человека, впрочем, не было подтверждено.
PP действует на рецептор Y4 в AP, NTS и дугообразном ядре, одновременно индуцируя расслабление желчного пузыря и ингибируя секрецию поджелудочной железы. Поскольку он действует как антагонист CCK, его работа приводит к задержке опорожнения желудка, что приводит к быстрому насыщению и уменьшению потребления пищи (Parker and Herzog, 1999; Balasubramaniam et al., 2006; Lin et al., 2009). Роль PP в подавлении аппетита доказывается исследованиями, демонстрирующими различие в концентрациях РР у людей в анорексическом состоянии и у людей в состоянии ожирения, когда концентрация увеличивается и уменьшается соответственно (Batterham et al., 2003b). Хотя РР является мощным подавителем аппетита, исследования, проведенные Clark et al. (1984), показали, что центральное введение РР стимулировало аппетит и приводило к увеличению потребления пищи. Данные по этому вопросу противоречивы. Необходимы дальнейшие исследования, чтобы определить направления воздействия РР на аппетит. Более того, исследования на пациентах с синдромом Прадера-Вилли и ожирением показали, что уровень PP после приема пищи у них снижен по сравнению со здоровой контрольной группой. Внутривенное введение РР у этих пациентов приводило к значительному снижению потребления пищи (Lassmann et al., 1980; Berntson et al., 1993). У мышей с РР-сверхэкспрессией наблюдается повышенная смертность в связи с сокращением потребления материнского молока (Kohno and Yada, 2012). Это подтверждает потенциальную эффективность РР в создании чувства насыщения. Кроме того, при использовании обинепитида (7TM Pharma) – мощного синтетического аналога PP и агониста рецептора Y4 – удалось добиться как снижения потребления пищи, так и потери веса (Davenport and Wright, 2014). Хотя его использование хорошо переносится с минимальными побочными эффектами, его испытания проводились только в течение 28 дней (Davenport and Wright, 2014). Следовательно, до того, как агонисты рецептора Y4 и агенты, полученные из РР, будут использованы в качестве средства для лечения ожирения, необходимы дальнейшие исследования для уточнения их долгосрочных эффектов.
Оксинтомодулин
Оксинтомодулин — это пептидный гормон, секретируемый в ответ на приём пищи. Как и GLP1, он синтезируется посредством посттрансляционной обработки препроглюкагонального пептида в эндокринных клетках кишечника и ЦНС (Cohen et al., 2003; Baggio et al., 2004; Habib et al., 2012). Его секреция происходит одновременно с GLP1 и PYY. Гормон достигает пика концентрации в течение 30 минут после приема пищи, после чего быстро разрушается DPPIV (Anini et al., 1999; Druce et al., 2009). Оксинтомодулин связывается с рецепторами GLP1 внутри ЖКТ, поджелудочной железы и дугообразного ядра, индуцируя снижение секреции желудочной кислоты и потребления пищи (Baggio et al., 2004; Dakin et al., 2004; Pocai et al., 2009). Центральное и периферическое введение оксинтомодулина увеличивает чувство сытости. Гормон снижает потребление пищи, что видно на моделях грызунов и человека, а также увеличивает затраты энергии (Dakin et al., 2002, 2004; Cohen et al., 2003; Baggio et al., 2004; Wynne et al. al., 2005). Кроме того, оксинтомодулин связывается с рецепторами глюкагона в поджелудочной железе, снижает концентрацию глюкозы в крови и способствует утилизации глюкозы (Maida et al., 2008). Хотя известно, что оксинтомодулин связывается с рецептором GLP1 и высвобождается одновременно с GLP1, точный механизм инкретинового действия оксинтомодулина при его связывании с рецептором глюкагона неизвестен (Pocai et al., 2009; Pocai, 2014). Гормон может выступать антагонистом глюкагона и индуцировать инсулинотропный эффект через местный, паракринный эффект, и/или он может активировать высшие центры мозга через гипоталамус. Поскольку сообщалось, что оксинтомодулин увеличивает затраты энергии, он может также активировать катехоламинергические и/или нейроны ПОМК, а также увеличивать активность волокон блуждающего нерва, иннервирующих бурый жир, тем самым увеличивая термогенез (Dakin et al., 2002; Wynne et al., 2006; Pocai, 2014). Хотя эти гипотезы правдоподобны, необходимы дальнейшие исследования механизмов действия оксинтомодулина до того, как его аналоги можно будет использовать при лечении ожирения.
Серотонин
Кишечный серотонин выделяется энтерохромафинными клетками ЖКТ в ответ на прием пищи. Серотонин оказывает свой эффект, действуя локально и системно на различные рецепторы 5-НТ (семейство рецепторов 5-НТ2 и рецептор 5-НТ4, экспрессированные на афферентных волокнах блуждающего нерва и других нейронах внутри ЦНС, а также клетках ЖКТ, сердца и надпочечников) (Halford and Harrold, 2012; Li et al., 2015; Stiedl et al., 2015). Аналоги серотонина, такие как лоркасерин, подавляют аппетит и уменьшают массу тела, а его антагонисты, напротив, увеличивают аппетит и массу тела (Halford et al., 1997; Savastano et al., 2007; Lam et al., 2008; Smith et al., 2010). Хотя серотонин уменьшает аппетит и способствует потере веса, действуя на ЦНС, обратные результаты были получены в эксперименте на животных: после перевода на западную диету они стали страдать ожирением, имея при этом повышенные концентрации серотонина (Crane et al., 2015). Кроме того, Crane et al. (2015) продемонстрировали, что ингибирование периферического синтеза серотонина препятствует развитию ожирения и метаболических нарушений. Это связано с тем, что серотонин ингибирует эффекты β-адренергических нейронов, иннервирующих бурую жировую ткань, и снижает термогенез. Аналоги серотонина были удалены с рынка из-за большого количества серьезных побочных эффектов, таких как психические расстройства, кардиотоксичность, наркомания и смерть (Onakpoya et al., 2016). Серотонин оказывает комплексное влияние на энергетический гомеостаз и пищевое поведение, оно требует более детального изучения и не будут рассмотрено в этой статье.
Эндоканнабиноидная система
Было показано, что биоактивные липиды эндоканнабиноидной системы, такие как анандамид, влияют на работу кишечно-мозговой оси. Эти молекулы синтезируются и секретируются внутри ЖКТ и действуют на эндоканнабиноидные рецепторы, в основном на каннабиноидные рецепторы 1 и 2 (CB1/CB2), которые являются GPCR-рецепторами в эндоканнабиноидной системе (Moran and Shanahan, 2014; Bauer et al., 2016). CB1 широко распространен в ЦНС и периферической нервной системе, есть в печени, поджелудочной железе и жировой ткани, тогда как CB2 преимущественно экспрессируется иммунными клетками в мозге, поджелудочной железе и жировой ткани (Mackie, 2008). Эндоканнабиноидная система регулирует различные физиологические функции, такие как перистальтика кишечника и аппетит. Это вызывает интерес, учитывая, что введение экзогенных каннабиноидов способствует ожирению (Mackie, 2008; Moran and Shanahan, 2014). Изучение антагонистов CB1, таких как римонабант и тарабант, показало, что они индуцируют потерю веса у индивидуумов, страдающих ожирением. Это подтверждает роль эндоканнабиоидной системы в регулировании аппетита (Christensen et al., 2007; Aronne et al., 2010) Cluny et al., 2011). Однако эти продукты были выведены с рынка, поскольку их применение приводило к тяжелым побочным эффектам, например, хронической депрессии (Aronne et al., 2010; Moran and Shanahan, 2014).
Ожирение сопровождается усиленной деятельностью эндоканнабиноидной системы, увеличением экспрессии CB1 и концентрации эндоканнабиноидов в плазме и жировых тканях (Izzo et al., 2009; Moran and Shanahan, 2014). Эти результаты подтверждаются исследованиями, демонстрирующими, что мыши с дефицитом CB1 устойчивы к ожирению при высокоуглеводной диете и обладают повышенной чувствительностью к лептину, который подавляет голод и увеличивает сытость (Ravinet Trillou et al., 2004; Cluny et al., 2011). Наконец, концентрация анандамида увеличивается во время голодания и вызывает голод за счет ингибирования экспрессирующих CB1 афферентных волокон блуждающего нерва. Это, соответственно, блокирует поступление сигналов в ЦНС и может приводить к снижению эффекта, производимого другими гормонами кишечника (Gómez et al., 2002; Kentish and Page, 2015). Хотя эта гипотеза еще не подтверждена, необходимо изучить роль эндоканнабиноидов в регуляции аппетита и энергетического гомеостаза, в особенности из-за связи эндоканнабиноидной системы со стрессом, памятью, иммунной функцией и настроением (Mackie, 2008). Факторы, влияющие на эти физиологические и психологические функции, могут быть связаны с пищевым поведением, усиливающимся при ожирении. Появились новые доказательства того, что эндоканнабиноиды (например, анандамид) связываются с рецептором TRPV1, который находится на мембранах большинства типов клеток в организме (Puente et al., 2011; Abdel-Salam, 2014). Активация TRPV1 с использованием низких доз поступающего с пищей капсаицина увеличивает термогенез, подавляет аппетит, улучшает функцию желудочно-кишечного тракта и усиливает потерю веса (Kawabata et al., 2009; Ludy et al., 2011; Ono et al., 2011; Abdel-Salam, 2014; Janssens et al., 2014). Существующий пробел в знаниях об эндоканнабиноидах и их связи с другими физиологическими рецепторами требует восполнения.
Расход энергии
Как было указано выше, ожирение развивается вследствие увеличения потребления энергии и снижения ее расхода. Сидячий образ жизни и малые затраты энергии способствуют набору веса (Grima и Dixon, 2013). Расход энергии связан не только с физической активностью, но и с термогенезом и поддержанием базального уровня метаболизма, хотя физическая активность может влиять на оба этих параметра (Melanson, 2017). Существуют различные исследования, свидетельствующие о том, что потеря жировой массы может происходить и без увеличения физической активности. Это доказывает, что затраты энергии могут возникать без системных физических упражнений, путем конкретного вмешательства (Panchal et al., 2012, 2013; Owen Bryn et al., 2014). Эти исследования бросают вызов предыдущей догме, где упражнения считались «золотым стандартом» регулирования энергозатрат и, следовательно, потери жировой ткани (Melanson, 2017). Кроме того, увеличение популярности фитнес-центров во многих западноевропейских странах и типичная схема их посещения также указывают на то, что физическая активность приводит к увеличению потери жира только в сочетании с правильной диетой (Австралия, 2009). Следовательно, нейрогуморальные сигналы, полученные из кишечника через кишечно-мозговую ось, могут активировать энергорегулирующие центры мозга ответ на потребление питательных веществ, модулируя расход энергии соответственно ее поступлению и тем самым способствуя благоприятному энергетическому балансу (Bauer et al., 2016).
Периферическое и центральное введение GLP1, оксинтомодулина и PYY приводит к увеличению затрат энергии за счет увеличения термогенеза и скорости обмена веществ (Dakin et al., 2002; Blouet and Schwartz, 2012). Blouet and Schwartz (2012) показали, что присутствие липидов в кишечнике активирует афферентные волокна блуждающего нерва, усиливая термогенез в бурой жировой ткани через CCK-зависимый путь. Это может служить доказательством существования оси «кишечник-мозг-бурый жир». Увеличивается и количество исследований, которые рассматривают вмешательство в работу этой оси для лечения ожирения (Blouet and Schwartz, 2012; Panchal et al., 2012, 2013; Brown et al., 2015 год). Например, поступление с пищей небольших доз капсаицина, который стимулирует активность кишечных афферентов блуждающего нерва, увеличивает выработку кишечных гормонов и тонус симпатической нервной системы, а также активирует бурый жир, что приводит к увеличению скорости обмена веществ и термогенеза, в итоге приводя к потере веса (Kawabata et al., 2009; Ludy et al., 2011; Ono et al., 2011; Abdel-Salam, 2014; Janssens et al., 2014). Исследование этой оси может помочь в лечении ожирения и связанных с ним сопутствующих заболеваний, открывая пути влияния на термогенез посредством терапии или диеты.
Кишечная микрофлора
Появляется все больше доказательств того, что микрофлора кишечника может влиять на ожирение и увеличение веса через несколько взаимозависимых путей. К ним относятся сбор энергии и последующее образование метаболитов (например, короткоцепочечных жирных кислот – SCFA), модификация поведения хозяина и влияние на воспалительные ответы в его организме (Moran and Shanahan, 2014).
Организм человека населяет более 3,9×1013 бактерий, большинство из них находится внутри ЖКТ и составляет сложную экосистему, называемую микрофлорой кишечника (Sender et al., 2016a, b). Исторически соотношение числа микробиальных и человеческих клеток оценивалось как 10:1, хотя последние исследования приводят соотношение 1,3:1 (Sender et al., 2016a, b). Эти микробы образуют сложную симбиотическую связь с хозяином, в которой хозяин обеспечивает питательную среду для микрофлоры, а микрофлора, в свою очередь, ответственна за метаболические, защитные и структурные функции, которые не кодируются геномом хозяина (Qin et al., 2010; Wang and Wang, 2016). Предполагается, что в микрофлору входит более 1000 видов бактерий (Схема 3), и их композиция изменяется на протяжении всей жизни. Это изменение вызвано зависит от широкого спектра факторов, например, диеты, использования антибиотиков, перенесенных болезней и даже механизма родов (Qin et al., 2010). Следовательно, состав микробиоты не статичен, а претерпевает динамические изменения.
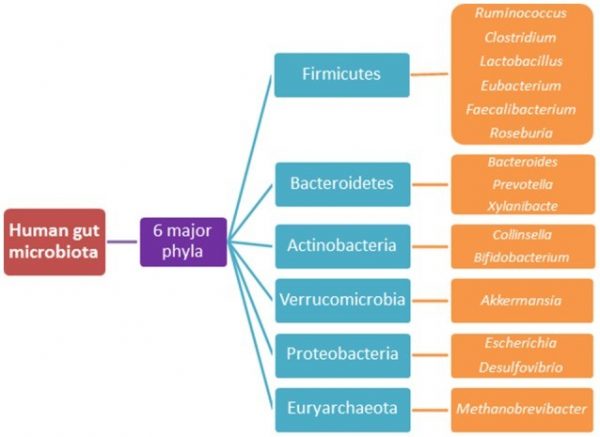
Рис. 3. 6 отделов кишечной микрофлоры и ее виды.
Микрофлора кишечника оказывает существенное влияние на метаболизм хозяина, что впервые было описано в исследовании Wostmann et al. (1983). Wostmann et al. (1983) продемонстрировали, что для мышей, лишенных микрофлоры, характерна меньшая степень ожирения, меньший уровень потребления энергии и получения энергии из стандартной диеты грызунов по сравнению с обычными мышами. Кроме того, Wostmann et al. (1983) собрали ряд исследований, анализировавших аномалии развития, возникающие у мышей, выращенных с микробиотой и без нее (таблица 3). Эти данные получили недавнее подтверждение в новых работах (Heijtz et al., 2011; Cho et al., 2012; Al-Asmakh and Zadjali, 2015). Это подчеркивает степень симбиотических взаимоотношений между хозяином и микрофлорой и тем, как изменение микрофлоры может влиять на состояние здоровья. Изучение эффектов, связанных с манипуляцией микробиотой, позволило бы лучше изучить физиологию хозяина по сравнению с использованием стерильных грызунов (Bauer et al., 2016). Это особенно актуально в отношении метаболических исследований, так как высокоуглеводное питание изменяет состав и разнообразие микрофлоры за короткий промежуток времени (David et al., 2014).
| Питание | Потребность в витаминах К и В Уменьшение жировой массы Нормальное или увеличенное потребление пищи |
| Баланс жидкости | Увеличение потребности в воде |
| Метаболизм | Уменьшение базального уровня метаболизма Повышение секреции аминокислот и мочи, незначительная экскреция уксусной кислоты Увеличение содержания мочевины и уменьшение содержания аммиака в кишечном содержимом Увеличение концентрации азота в содержимом слепой кишки и фекалиях Изменение ответа на анестетики |
| Кровообращение | Уменьшение общего объема крови Снижение кровотока в коже, печени, легких и ЖКТ Повышение концентрации холестерина и количества эритроцитов, увеличение гематокрита |
| Печень | Уменьшение размера Повышение концентрации ферритина и холестерина |
| Легкие | Истончение альвеолярных и капсульных стенок, уменьшение числа ретикулоэндотелиальных элементов |
| Морфология кишечника | Уменьшение общей массы кишечника Уменьшение общей площади поверхности тонкого кишечника Укорочение ворсинок повздошной кишки и удлинение дуоденальных ворсинок Укорочение крипт тонкого кишечника Истончение собственной пластинки тонкого кишечника, уменьшение числа клеток и замедление их регенерации Уллинение слепой кишки, истончение ее стенки |
| Моторика кишечника | Увеличение массы мышечной ткани с удлиненными и гипертрофированными миоцитами в слепой кишке Замедление прохождения пищевого комка |
| Кишечные функции | Улучшение абсорбции витаминов и минералов, изменение поглощения других макромолекул Изменение содержания ферментов, повышение концентрации трипсина, химотрипсина, инвертазы и муцина Уменьшение содержания жирных кислот и отсутсвие жирных кислот с циклической или разветвленной цепью в кишечном содержимом, экскреция преимущественно ненасыщенных жирных кислот |
| Эндокринная функция | Уменьшение потребления йода в щитовидной железе Снижение двигательной активности и гиперчувствительность к адреналину, норадреналину и вазопрессину |
| Электролитический статус | Защелачивание кишечной среды Увеличение концентрации кальция и цитрата, незначительное количество фосфата в моче Снижение концентрации натрия и хлорида в кишечном содержимом |
De Filippo et al. (2010) сравнили и охарактеризовали различия между здоровыми детьми, придерживающимися западной и сельской диеты, и выявили четкую связь между составом микрофлоры и потребляемыми пищевыми продуктами. Хотя De Filippo et al. (2010) исследовали детей из Италии, которые придерживались западной диеты, исследование не предусматривало, что эти дети могли быть приучены к средиземноморской диете, которая считается золотым стандартом здорового питания (Sánchez-Villegas et al., 2016). Ясно то, что существуют различия между двумя культурами, потребляемой пищей и, следовательно, составом микрофлоры. Это исследование, как и другие, показало, что западная диета и ожирение связаны с повышенным количством бактерий типа Firmicutes по отношению к количеству бактерий типа Bacteroidetes. Этот баланс изменяется в противоположную сторону при хирургических и диетических вмешательствах (Ley et al., 2006; Turnbaugh et al., 2009; De Filippo et al., 2010; Furet et al., 2010). Возможность использования соотношения Firmicute-Bacertoidetes для определения фенотипа ожирения активно обсуждается, поскольку более поздние исследования не подтвердили эту гипотезу (Zhang et al., 2009; Schwiertz et al., 2010; Finucane et al., 2014). Различия на уровне рода и вида бактерий по сравнению с уровнем отдела могут быть связаны с изменениями в метаболизме (Bauer et al., 2016).
Duca et al. (2014) показали, что у крыс, выращенных без контакта с микробами, развиваются значительные видовые различия после трансплантации микрофлоры крыс, страдающих ожирением, или микрофлоры людей с ожирением. Duca et al. (2014) выявили 25 оперативных таксономических единиц (ОТЕ) у страдающих ожирением доноров и реципиентов, которых не было у устойчивых к ожирению доноров и реципиентов. Эти дополнительные 25 ОТЕ обладали способностью извлекать дополнительную энергию из рациона. Это исследование подтверждает результаты более ранних работ, в которых предполагалась связь между ожирением и микрофлорой, богатой генами, ответственными за производство ферментов, собирающих энергию из неусвояемых углеводов (Turnbaugh et al., 2009; Duca et al., 2014). Связь между составом микрофлоры, сбором энергии и ожирением более сложна, чем предполагалось. Исследования показали, что производство энергии и производство метаболитов в форме короткоцепочечных жирных кислот не коррелируют с увеличением веса, и что некоторые КЖК могут обладать полезной ролью в отношении метаболизма хозяина и регуляции энергии (Tims et al., 2013; Bauer et al., 2016).